Friday, May 29, 2015
Tuesday, February 04, 2014
Thursday, January 30, 2014
Raising (a little) money for Cancer Research
As much as I like science (doing science or reading about it) I also like running. Luckily running and thinking are quite complementary activities so I do not see the time I spend running as something detracting from my work. Since I arrived to +Moffitt Cancer Center in the summer of 2008 I have participated in a charity race called Miles 4 Moffitt.
 |
| Miles for Moffitt |
This event, organised for +Moffitt Cancer Center, raises money for local cancer researchers. I know a few of my colleagues that applied and received some of that money and I know how much it helps junior researchers at Moffitt to get a little bit of funding so they can test new ideas that would go untested otherwise. This year I am not only running but also trying to raise a very modest sum of money for this cause.
 |
| Yours truly on the left with one of the race t-shirts |
Yours truly: http://www.kintera.org/faf/donorReg/donorPledge.asp?ievent=1088017&supId=400183943&extSiteType=4
Some of my friends and colleagues at Moffitt's IMO are also running and they could do with your help as well:
And that's all really. I am hoping to update the list very soon. I am optimistic about +Philip Gerlee, +Jacob Scott, +Jill Gallaher and maybe +Arturo Araujo ?
Friday, January 17, 2014
Get it while it is still fresh! A mathematical model of stem cell-driven tumourigenesis
A couple of months ago +Jacob Scott and I (as well as +Anita Hjelmeland , +Prakash Chinaiyan and +Alexander Anderson ) got our work accepted in PLOS Computational Biology and finally it is available online here.

This is an example of a simulation where on the left you can see the different types of cells (stem in red and non stem in green and blue) as well as blood vessels; whereas on the right you can see the concentration of oxygen (from white where there is abundance to red where there is hypoxia). The work is available to anybody since it is a PLOS paper. +Jacob Scott has also produced a nice description on his blog here. So go ahead and take a look if you are interested in mathematical oncology, cancer stem cells, both or either.
Also thanks to +Alexander Anderson for this paper. This work started a few years ago when I was a postdoc at his group using a mathematical tool, the hybrid discrete-continuum cellular automaton, that I learned from him. Nonetheless he let me take responsibility for the project while at the same time contributing to it with his expertise and ideas.
Expect to see new results from this model soon, it is difficult to stop +Jacob Scott when he has an idea and I am afraid he has quite a few involving versions of this model.
Monday, January 13, 2014
Heterogeneity in CML
Followers of this blog (or even just casual readers) know that heterogeneity is a key aspect of cancer. Not that I am saying that this is my idea, far from it. Many people have championed it in the last few years including my colleague +Alexander Anderson. A couple of years ago CRUK's Charles Swanton and his team produced convincing clinical evidence of the existence of phenotypic heterogeneity in kidney cancer [news,article] and a lot more people started paying attention. Since then researchers have found evidence of heterogeneity in other types of cancer such as prostate, bone or Barrett's esophagus.
Why is this important you say (or not...)? Because we are moving towards the use of targeted therapies. Therapies that, for the most part, assume that there are critical *targetable* mutations that all tumour cells share. Sadly this is unlikely to be true for most cancers.
An exception could be CML or Chronic Myeloid Leukemia, a type of tumour in which I started working recently during the IMO workshop (+Artem Kaznatcheev describes it nicely here [link]). Our clinical experts were quite clear that there is no heterogeneity in CML. There is only a key mutation, BCR-ABL, driving CML that if messed with, controls the cancer. That lack of genetic heterogeneity could explain why treatments like imatinib are so effective.
But it does not work every time, it does not work the same for everybody and even if there is not substantial genetic heterogeneity there are other elements that explain intra-tumour heterogeneity. +Chandler Gatenbee and I came with this list, which is certainly not exhaustive, during a brainstorming session:

Why is this important you say (or not...)? Because we are moving towards the use of targeted therapies. Therapies that, for the most part, assume that there are critical *targetable* mutations that all tumour cells share. Sadly this is unlikely to be true for most cancers.
An exception could be CML or Chronic Myeloid Leukemia, a type of tumour in which I started working recently during the IMO workshop (+Artem Kaznatcheev describes it nicely here [link]). Our clinical experts were quite clear that there is no heterogeneity in CML. There is only a key mutation, BCR-ABL, driving CML that if messed with, controls the cancer. That lack of genetic heterogeneity could explain why treatments like imatinib are so effective.
But it does not work every time, it does not work the same for everybody and even if there is not substantial genetic heterogeneity there are other elements that explain intra-tumour heterogeneity. +Chandler Gatenbee and I came with this list, which is certainly not exhaustive, during a brainstorming session:
There are tumour cells with different degrees of stemness, cells at different states of the cell cycle, different proliferative potential, expresion of Beta-catenin...and that is before we even start considering the microenvironment of the tumour (access to oxygen, other cells, that is, non-tumour cells...). Could it be the reason why not all patients respond the same way to the, otherwise very successful imatinib? I think there is a good chance that heterogeneity could be behind that. Let's now see if our clinicians at Moffitt (or maybe elsewhere) can give us a information we could use to correlate CML heterogeneity and response to imatinib.
Wednesday, December 18, 2013
Can (computational) models be trusted?
As a computational modeller I am part of a group of people doing science in a way that was impossible only a few decades ago. A lot of computational modelling combines some of the features of theoretical work (finding out the essential elements of the reality that needs to be captured and creating a computational representation of it) with experimental work (using the computer model as surrogate of the reality to be studied to quickly and exhaustively use it for experiments). Here is an example where Ziv Frankenstein (working with +Alexander Anderson , +Simon Hayward , +Gus Ay and myself) has captured what we thought were some of the essential cellular and microenvironmental players in prostate cancer progression and used that to study how the microenvironment of the tumour can help explain how the tumour evolves.


This week Aeon magazine published this piece by Jon Turney where he asks whether we should trust computational models at all. Computational models allow us to explore very complicated scenarios that would be impossible to study otherwise. The issue the article raises is whether we are beginning to rely too much on these computer models. This is a genuine concern, in some fields (I think +Artem Kaznatcheev might agree with me that social sciences could be one of those) experiments are really hard and the data scarce. This means that the computational models will have to be either too abstract (limiting the detail predictive power of the model) or risk having to make too many assumptions about aspects that are not clear (and thus leading to wrong predictions).
Bottom line: computational models are extraordinarily useful but depend on having good data and good understanding of what is being studied. We are much better of for having the ability to use them but be careful of detailed predictions when little is known for they are likely to be just guesses.
Wednesday, November 27, 2013
Introducing mathematical oncology to cancer biology and medical students
This Wednesday, as I did last year ago, I am helping with a lecture series entitled Modern Basic Tools of Research at the Moffitt Cancer Center. Last year's was a pleasurable experience and for 2h we talked about different ways to model growth in tumours. First with growth laws describing population change over time (examples of those laws can be found here), then with more mechanistic models where the dynamics of the tumour growth emerge from the way that different cells interact with each other and with their environment.

This year going that far will be more difficult: my 2h have been reduced to 30m. Thus I am trying to change the focus of the lecture and maybe limit my ambitions. Interestingly this could be a blessing in disguise since I might be forced to try to figure out what is the essence of mathematical modelling of cancer and explain that to smart people that do not have a background (or maybe not even an interesting) in mathematical modelling.
And why 30m this year? Because due to time constraints, the same 2h lecture slot will be used to teach about biostatistics, bioinformatics and mathematical modelling. And that is good: since I have been working in this field every biologist and medical professional I met expected me to be a statistician after describing myself as a mathematical oncologist. This is going to be a great opportunity to explain what we have in common but also the ways in which we work differently.
Hoping to not misrepresent excessively what biostastisticians and bioinformaticians do (expect a line saying UPDATE at the end of this post very soon!), their work is incredibly useful to both cancer biologists and doctors since it allows them to figure out what the data says about a specific biological process or clinical trend. It also allows them to know whether that trend or pattern is meaningful or not or whether they have collected enough experimental points or clinical data to make any statement about it.
The best part is that all that extra information usually comes at very little cost to the experimentalists. Mathematical oncologists on the other hand, we tend to be somewhat more difficult to work with since we need to have an understanding of the biological mechanisms underpinning the cancer we are studying. We, for instance, take a look at the diagram after the first paragraph and think: do all these cell interact with each other? if so how? these tumour cells, are they all the same? where do they come from? do they come alone or together? do they usually arrive in the neighbourhood of the other cells in the diagram? if not, do they sit and wait? Maybe some of the questions would be part of the conversation between experimentalists and but many of them seem to arise when mathematical and computational modellers are involved. When implementing these ideas into a computer, ambiguities are not an option. Ideas that might work in your mind or mine come crashing down when subjected to the cold logic of a computer.
It takes time.
The advantage? We can test new hypotheses, generate novel ones, get molecular, clinical, cellular data and integrate it into the model, we can get all those single cell level measuraments and feed them directly into the model, we can take all these population level experiments and figure out what hypothesis explain them better. We can use that to understand the biology of the cancer, to design new biological experiments, to predict better clinical treatments, to hypothethise how new ones would impact patients in the clinic.

This year going that far will be more difficult: my 2h have been reduced to 30m. Thus I am trying to change the focus of the lecture and maybe limit my ambitions. Interestingly this could be a blessing in disguise since I might be forced to try to figure out what is the essence of mathematical modelling of cancer and explain that to smart people that do not have a background (or maybe not even an interesting) in mathematical modelling.
And why 30m this year? Because due to time constraints, the same 2h lecture slot will be used to teach about biostatistics, bioinformatics and mathematical modelling. And that is good: since I have been working in this field every biologist and medical professional I met expected me to be a statistician after describing myself as a mathematical oncologist. This is going to be a great opportunity to explain what we have in common but also the ways in which we work differently.
Hoping to not misrepresent excessively what biostastisticians and bioinformaticians do (expect a line saying UPDATE at the end of this post very soon!), their work is incredibly useful to both cancer biologists and doctors since it allows them to figure out what the data says about a specific biological process or clinical trend. It also allows them to know whether that trend or pattern is meaningful or not or whether they have collected enough experimental points or clinical data to make any statement about it.
The best part is that all that extra information usually comes at very little cost to the experimentalists. Mathematical oncologists on the other hand, we tend to be somewhat more difficult to work with since we need to have an understanding of the biological mechanisms underpinning the cancer we are studying. We, for instance, take a look at the diagram after the first paragraph and think: do all these cell interact with each other? if so how? these tumour cells, are they all the same? where do they come from? do they come alone or together? do they usually arrive in the neighbourhood of the other cells in the diagram? if not, do they sit and wait? Maybe some of the questions would be part of the conversation between experimentalists and but many of them seem to arise when mathematical and computational modellers are involved. When implementing these ideas into a computer, ambiguities are not an option. Ideas that might work in your mind or mine come crashing down when subjected to the cold logic of a computer.
It takes time.
The advantage? We can test new hypotheses, generate novel ones, get molecular, clinical, cellular data and integrate it into the model, we can get all those single cell level measuraments and feed them directly into the model, we can take all these population level experiments and figure out what hypothesis explain them better. We can use that to understand the biology of the cancer, to design new biological experiments, to predict better clinical treatments, to hypothethise how new ones would impact patients in the clinic.
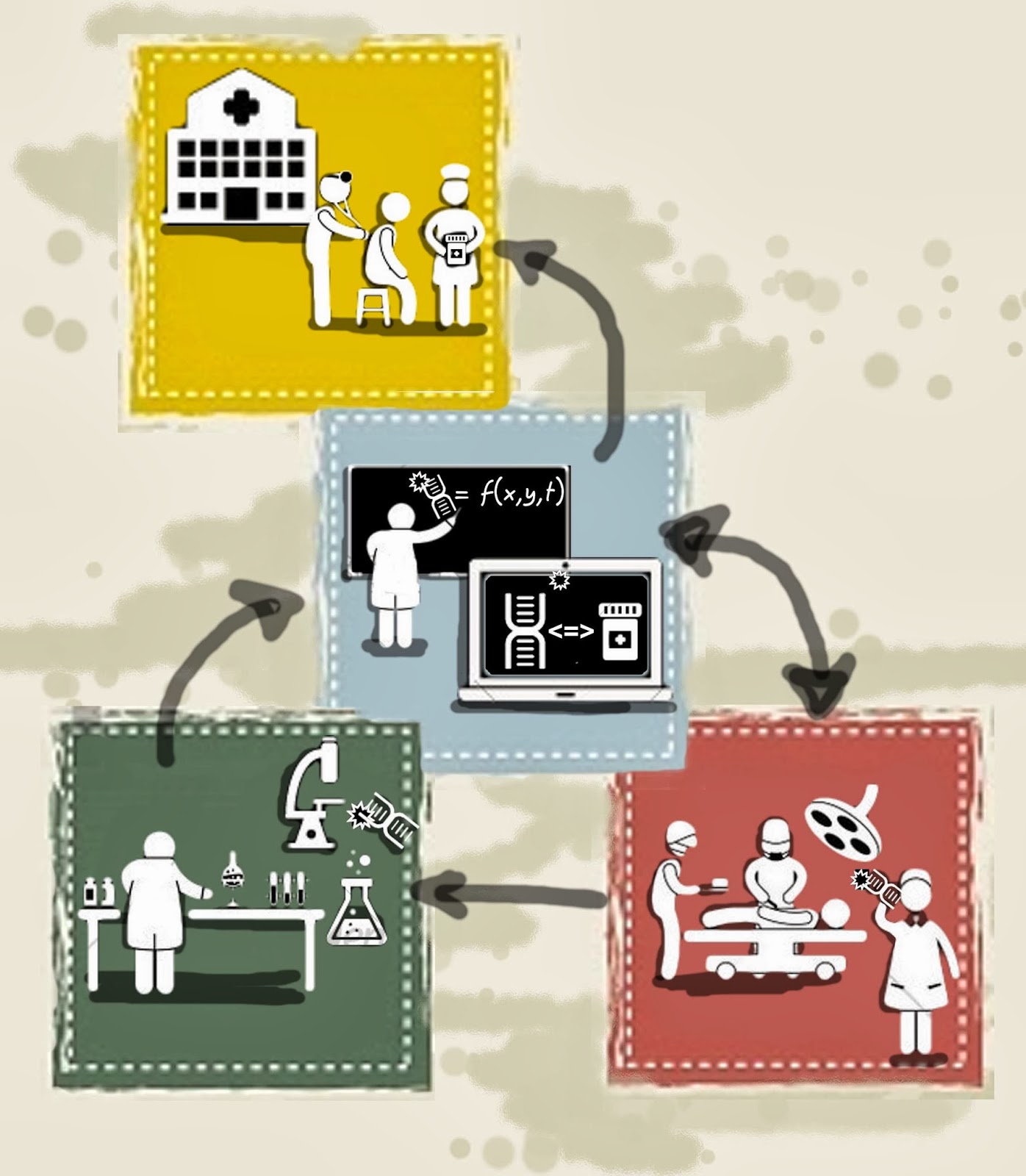
If you are an experimentalist you should know that using mathematical model will require you to work in a different way, to ask different questions and to view of your research with different lenses but it is worth it.
By the way, all the figures in the post have been crafted by our very own +Arturo Araujo .
Monday, September 02, 2013
Pint of Science
+Arturo Araujo and myself think that our mission as scientists consists, not only in producing new and interesting discoveries, not only contributing to research a cure to cancer but also in making sure that the society at large understands the nature of what we do and the implications of our work.

Is for that reason that, together with +Angela Rey, we jumped into the opportunity to help +Parmvir Bahia start +Pint of Science US , and offshoot of the UK based Pint of Science, but with a twist: that we will produce regular meetings and podcasts where people in Tampa and beyond will have the chance to interact with our local scientists.
The first of these events will take place tomorrow at The New World Brewery in Ybor City and will feature a friend and colleague at +Moffitt Cancer Center 's IMO: +Jacob Scott. So science, medicine and mathematics will be the ingredients for tomorrow's event at 7pm (9/3/2013). Come if you can, more details at Pint of Science US.
Monday, July 29, 2013
The story of a paper
Together with my good friend and collaborator +Jacob Scott and our new collaborator +Artem Kaznatcheev , we have recently wrote (what I hope you will find) an interesting paper where we explored one of older games (emergence of motility in tumours mostly made of rapidly proliferating cells) and studied what happened when cells grow and reach an edge or a boundary. Normally I would try to recapitulate how did we do our work and what the results are but this time I do not need to bother since my collaborators are so much faster and better than me at this.
For a story of how this work started and how we did this work take a look at +Jacob Scott's latest post.
The work itself has been described in detail by +Artem Kaznatcheev on his own blog in the following post.

For a story of how this work started and how we did this work take a look at +Jacob Scott's latest post.
The work itself has been described in detail by +Artem Kaznatcheev on his own blog in the following post.

Friday, July 26, 2013
Writing grants
Unfortunately, a good deal of the work of a professional biomedical scientist in the US involves writing grants. It might provide a consolation to some to know that almost 100 years ago scientists still had to write grant proposals. I am not sure how many would have looked like this though:


Subscribe to:
Posts (Atom)